New Ag International SEPT/OCT 2020
Nickel (Ni) is a structural component of the enzyme urease, making it essential for monocot and dicot plants. This fact, together with its irreplaceable role for ordinary functioning of hydrogenase and six other enzymes – some of which are central in nitrogen metabolism – have justified its inclusion, albeit rather recently, in the exclusive group of essential plant nutrients. Highest Ni-deficiency sensitivity is found in field-grown, nitrogen-fixing plants, and in plants utilizing urea as their primary nitrogen source.
Normally, plant leaves contain Ni at 0.1–5ppm in dry matter, which maintains plants' optimal growth and productivity. If Ni deficiency occurs, it will likely be associated with soils having pH>6.7, or after excessive applications of zinc (Zn), copper (Cu), manganese (Mn), iron (Fe), calcium (Ca) and/or magnesium (Mg). In most plant species, Ni deficiency appears as leaf-tip necrosis, and/or as general chlorosis, attributable to general nitrogen (N) deficiency. The easiest and most effective method for correcting acute Ni deficiency is foliar spraying with a dilute solution of NiSO4, or another water-soluble Ni fertilizer. At supra-optimal concentrations, Ni2+ becomes phytotoxic, mainly due to over-generation of reactive oxygen species (ROS), which jeopardizes the function of many enzymes and proteins, causing subsequent impairment of photosynthesis, source-sink relationships and reducing crops productivity.
Variations in Ni levels Nickel is a heavy metal, rather rare in earth’s crust. It is chemically similar to Fe, cobalt (Co), Cu and Zn. Nickel is present in most agricultural soils, commonly at 20-30ppm, and seldom exceeding 50ppm. But Ni concentration can exceed 10,000ppm, where soil's parent materials is high in Ni, like the serpentine group of minerals, and goethite and hematite. Nickel concentrations can also be higher due to atmospheric deposition, near metal refineries, and owing to soil application of biosolids and sewage sludge. Similar to Zn2+, Cu2+, Fe2+ and Co2+, the most important soil parameter affecting Ni solubility and plant availability is soil pH, whereby pH>7.5 enhances its oxidations which, in turn, markedly compromises its plant availability. High soil organic matter readily adsorbs and complexes Ni and reduces its plant availability as well. Also, high soil concentrations of any of the above-mentioned divalent cations can inhibit Ni uptake, due to their mutual competition for root uptake sites.
Other factors reducing plants' Ni uptake are cool and/or dry soil conditions, and nematodes damage to feeder roots. However, applying elemental sulphur to the soil increases the solubility and uptake availability of all these nutrients, due to the acidifying effect of sulphur when it is oxidized and hydrated. Soil testing for Ni as a plant nutrient is not a routine practice, since agronomic know-how regarding its requirements is insufficient. Using the DTPA soil test is the most effective extractor for assessing its contents in most soils. However, standards for defining soils as Ni-deficient or Ni-sufficient have not yet been established.
Central role of Ni in plants' N metabolism Plants requirement for nickel is the lowest among all essential mineral nutrients, and this might be the fact behind our relatively meagre knowledge about plants' Ni nutrition. But let's illuminate the issues that have been established so far. Plants take it up in the form of soluble Ni2+, which is also its preferred oxidation state in biological systems and where it rarely exists, as Ni+, and Ni3+. Nickel forms stable complexes, with histidine, cysteine, citrate and malate. It is readily mobile within plants, so it is likely that Ni deficiency symptoms should show on older leaves. In some species, it's the developing seeds that feature main Ni sinks.
Ni concentration in most species' leaves normally ranges 0.1-5ppm in dry matter, but can be highly variable, depending on its availability in soils, plant species and season. Tissue Ni concentrations above 10ppm are considered phytotoxic in sensitive crop species, and above 50ppm in moderately tolerant species. Pecan tree's sufficiency range is 2.5-30ppm, and toxicity occurs at concentrations above 100ppm. This crop is, hence, considered a high Ni-requiring species. But there are some 350 Ni “hyperaccumulating” species, which gather 1,000ppm Ni without phytotoxicity symptoms.
Nickel participates in the structure and functioning of at least eight enzymes, one of which is [NiFe]-hydrogenase. It recycles the hydrogen produced by the nitrogenase enzyme in cells fixing atmospheric N2. But it is the enzyme urease that has justifiably become the focus of most Ni-related studies. Urease, whether produced by plants, microbes or animals, is always dependent on Ni availability for its normal activity of converting urea to ammonium (NH4+), which plants can use as an N feedstock.
Jack-bean urease contains 12 Ni atoms that are crucial for the catalytic function of this enzyme. In other plants, Ni works as a cofactor to enable urease functioning. These make Ni a cornerstone in normal N nutrition of plants. No wonder, therefore, that when urea was supplied as a sole N source to duckweed, tobacco, zucchini, tomato, rice or oilseed rape, in absence of Ni, plants growth was markedly reduced.
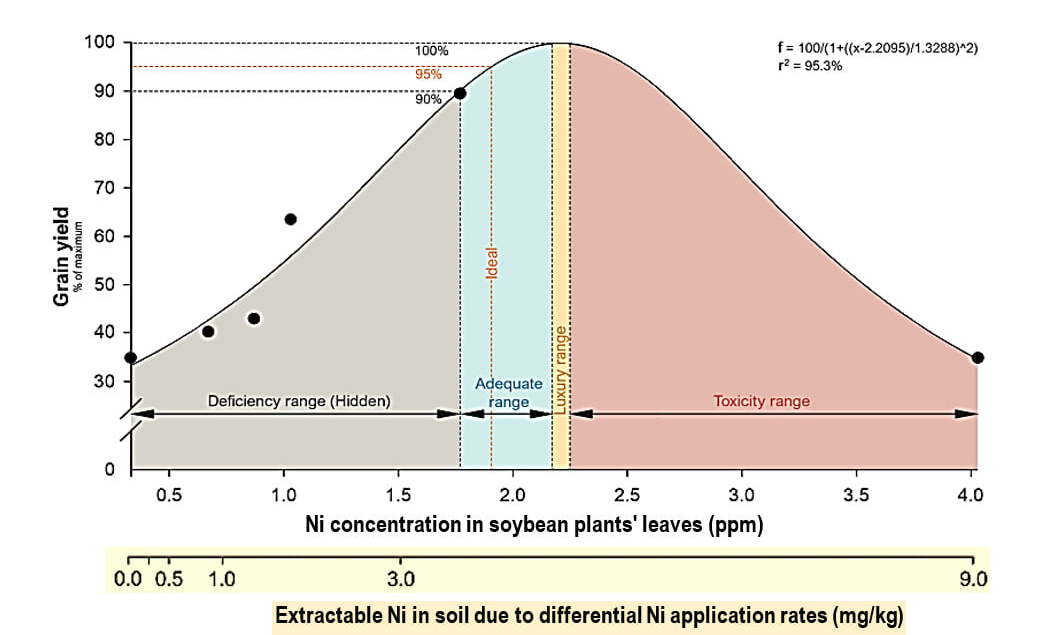
When soybean plants are deprived of Ni, they accumulate urea at toxic concentrations, up to 2.5 percent, which provokes necrotic lesions on their leaflet tips. This phenomenon occurs equally if N's origin is mineral or obtained by N fixation (Eskew et al. 1983). But soil- or foliar-applied Ni quickly restores plants' urease activity, decreasing urea concentrations and reestablishing normal growth. Similarly, Freitas et al. (2019) showed that judicious Ni application remarkably increased N metabolism in soybean plants, owing to higher nodules' nitrogenase activity, accompanied by greater Ni concentrations in areas of highest metabolic activity, thereby indicating a former Ni deficiency in the plant. He could draw a Ni optimum response curve to differential Ni soil-application rates. Figure 1 shows the dependence of soybean grain yield, and its concentration in the plants leaves, on Ni application rate.
Figure 1. The effects of Ni application rate on its concentration in soybean plants' leaves, and on their grain yield. Source: Freitas et al 2019
Defensive role Ni is also involved in the synthesis of phytoalexins that plants produce to defend themselves against pathogens. An additional protection mechanism related to Ni sufficiency is increased lignin production, which strengthens plants' cell walls, thereby contributing to disease resistance.
Chlorosis and leaf-tips necrosis are reliable Ni deficiency symptomsUrea accumulation in leaf-tips of monocot and dicot plants can serve for detecting Ni deficiency. Seedlings of Gramineous species, e.g. wheat, barley and oat, grown at Ni deficiency, accumulate urea and show leaves' interveinal chlorosis, severe leaf-tip necrosis and also fail to unfold. This deficiency also results in significantly reduced root and shoot growth. Ni deficiency also markedly boosts plant senescence and reduces tissue Fe contents. Owing to its major role in plants' nitrogen nutrition, Ni deficiency is sometimes expressed as overall chlorosis, similar to general N deficiency. Disturbed N metabolism in Ni-deficient plants also exhibit marked changes from the norms of concentrations of organic acids and other solutes. These phenomena reveal that urea is a common component in N metabolism, and that its concentration has to be maintained at a low level in order to prevent phytotoxicity. In pecan trees, Ni deficiency markedly reduces leaflets size and results in their discoloration.
Correcting Ni deficiency is rather simple Crops responding to foliar and soil Ni applications are normally those that utilize ureides (acyl derivatives of urea) as the main internal transference form of N. The clearest agronomic responses to Ni have been observed in N2-fixing plants, like soybean, beans, pea and other legumes. Ni deprivation of these crops results in delayed nodulation and reduced root and shoot growth. The addition of actual Ni at 1 microgram/L to the nutrient media of these crops is sufficient to prevent the deficiency damages and symptoms mentioned earlier. Ni requirement of most annual plants can be satisfied by applying around 0.56kg/ha (0.5lbs/acre) of actual Ni. Application of Ni fertilizers might be needed at the following growth conditions: i) urea is the primary N carrier; ii) leguminous crops; iii) the soil is poor in mineral content and/or has a pH>6.7; iv) high rates of Zn, Cu, Mn, Fe, Ca or Mg have been applied in previous years.
Soluble nickel salts, like Ni-sulphate, Ni-chloride or Ni-nitrate, or a nickel chelate, are suitable for preventing or correcting plant Ni deficiency (Table 1). They can be soil-applied or fertigated at 0.56kg/ha, or foliar-sprayed at 0.03–0.06ppm of actual Ni. Also, municipal biosolids are a good source of nutritious Ni.
Ref.: Liu et al. Florida, 2017
Fertilizer
Formula
Ni (%)
Anhydrous nickel sulphate
NiSO4
37.5
Nickel chloride
NiCl2 •6H2O
37.2
Nickel sulphate ("nickelous sulphate")
NiSO4 •6H2O
32.1
Nickel nitrate
Ni(NO3)2 •6H2O
20.2
Nickel(II) EDTA complex
Ni-C10H16N2O8
16.7
Nickel Plus*
Complex
5.4
Sewage sludge
Composite
2.4–5.3
'* Contains 5%N and 3%S
Ni toxicity is not a lesser problem than Ni deficiency Generally, Ni toxicity is more abundant than Ni deficiency. The main reason for excessive Ni contents in plants is disproportionate Ni in their environment. Extreme plants' Ni commonly occurs when the local soil has evolved from a Ni-rich mother rock, or the irrigation water passes via such minerals, and due to anthropogenic activities, like metal mining, metal smelting, electroplating, vehicle emissions, fossil fuel burning, disposal of household, municipal and industrial wastes, or soil application of high-Ni sewage sludge.
Mineral fertilizers, particularly phosphatic ones, have variable levels of Ni2+. Polluted soils may exhibit Ni2+ concentrations in the range of 200-26,000 mg/kg, which are 10-850 fold higher than the normal range.
Critical toxicity levels in crop plants are in the range of >10µg/gDW in sensitive species, to >50µg/gDW in moderately tolerant species. Sachan & Lal (2017) showed that high concentrations of Ni2+ may result in deficiency of essential cations like potassium (K), Zn, Mn and Cu. High affinity of Ni2+ to sulfhydryl (-SH) groups and disulfide bonds, may damage the secondary structure of proteins and the activity of metabolic enzymes. Enzyme activity is also compromised in the presence of high Ni2+ due to competitive binding with their central metal ion. Excess Ni2+ adversely affects seed germination by hampering the activity of hydrolytic enzymes, such as amylases and proteases. During germination, excess Ni2+ also inhibits all energy-dependent cellular processes, resulting in slow radicle and plumule emergence. Several plants, including maize, have displayed inhibited lateral root formation and development, due to Ni2+ toxicity. Ni2+ was also found to negatively affect photosynthesis mainly by inhibiting important enzymes involved in chlorophyll biosynthesis, such as ALA dehydratase and protochlorophyllide reductase. Excessive Ni2+ also affects plants cell division, exerting a mutagenic effect. At higher concentrations, Ni2+ leads to oxidative stress in plants by increasing the amount of hydroxyl radicals, superoxide anions, nitric oxide and hydrogen peroxide.
However, the use of Ni-hyperaccumulator plants has shown its effectiveness in remediating Ni-contaminated soils employing an agromining process (Nkrumah et al. 2019).